

Fakten über Superoxid-Dismutasen: Rolle in der Redox-Signalisierung aufgedeckt
Die SODs stellen das erste enzymatische Abwehrsystem gegen die radikale Schädigung durch Sauerstoff dar: daher ist dieses Enzym für alle aeroben Organismen essentiell, aber nicht für Anaerobier. Zur Unterstützung dieser Hypothese glaubte McCord, dass die Existenz eines aeroben Organismus hauptsächlich von seiner Fähigkeit abhängt, SODs zu produzieren, da ihr Mangel für die Sauerstoffempfindlichkeit verantwortlich ist und ein Überleben nur in einer anaeroben Umgebung ermöglicht.
Unter physiologischen Bedingungen halten die Superoxiddismutasen zusammen mit den nicht-enzymatischen ROS-Fängern wie den Vitaminen E, A und C einen stabilen Zustand zwischen oxidativen und antioxidativen Systemen aufrecht (Russo et al., 2011). Die Dysregulation der Redox-Homöostase, die durch ein Ungleichgewicht zwischen ROS-Produktion und Abfangkapazität bestimmt wird, bestimmt erhebliche zelluläre Schäden wie Membranlipoperoxidation, Nukleinsäure- und Strukturveränderungen von Proteinen, die zu neurodegenerativen und kardiovaskulären Erkrankungen beitragen.
In den letzten Jahren haben viele Daten aus in vitro-Studien, die an vielen Zelllinien, hauptsächlich an Neuroblastomzellen SK-N-BE, durchgeführt wurden, gezeigt, dass SOD1 sezerniert wird und in der Lage ist, über den muskarinischen M1-Rezeptor zelluläre Signalwege zu aktivieren, an denen ERK1/2- und AKT-Aktivierung beteiligt sind; diese Effekte sind mit einem intrazellulären Kalziumanstieg verbunden, der noch verstärkt wird, wenn diese Zellen mit mutiertem SOD1G93A stimuliert werden.
Die intrazelluläre zytosolische Lokalisation von SOD1 ist umstritten. Jüngste Nachweise, die in transfizierten Neuroblastomzellen der Maus (Neuro2) durchgeführt wurden, zeigten, dass sowohl Wildtyp-SOD1 (wt-SOD1) als auch SOD1-Mutanten in luminalen Strukturen des endoplasmatischen und Golgi-Apparats verteilt sind (Urushitani et al., 2008). Die ersten experimentellen Hinweise darauf, dass einige Zelllinien in der Lage sein könnten, die Cu,Zn-Superoxiddismutase zu sezernieren, reichen viele Jahre zurück, als wir zum ersten Mal die Sekretion dieses Proteins durch Experimente in Hepatozyten und Fibroblasten (Mondola et al., 1996), Neuroblastom SK-N-BE-Zellen (Mondola et al., 1998; Gomes et al., 2007; Polazzi et al., 2013) und in Epithelzellen aus dem Thymus (Cimini et al., 2002) zeigten.
Darüber hinaus konnten wir zeigen, dass in humanen Neuroblastom SK-N-BE-Zellen, die aufgrund einer erhöhten Empfindlichkeit gegenüber ROS (Shutt et al., 2010) eine größere Empfindlichkeit gegenüber Glukose-Entzug-induzierter Zytotoxizität aufweisen, der SOD1-Export unter normalen Bedingungen stattfindet und nach oxidativem Stress erhöht ist (Mondola et al., 1996, 1998). In der Folge konnten wir zeigen, dass SOD1 in humanen Neuroblastomzellen SK-N-BE über einen mikrovesikulären sekretorischen Weg exportiert wird, der durch Brefeldin-A (BFA) sowie durch 2-Desoxyglucose und Natriumazid, das den intrazellulären ATP-Pool reduziert, beeinträchtigt wird (Mondola et al., 2003).
Hinweise auf die Rolle der Mangan-Superoxid-Dismutase in ... - Hindawi You Need To Know
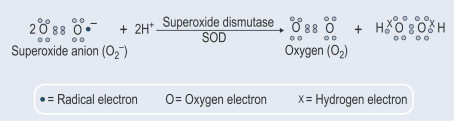

Ein weiterer wichtiger Aspekt war die Entdeckung, dass neben dem konstitutiven SOD1-Export auch die Sekretion dieses Enzyms induziert wird. In diesem Zusammenhang zeigten wir (Santillo et al., 2007), dass SOD1 aktiv aus Synaptosomen des Rattenhirns sowie aus hypophysären GH3-Zellen der Ratte freigesetzt wird, die ein gutes Modell zur Untersuchung der induzierbaren SOD1-Freisetzung darstellen, da sie die gesamte neuronale Proteinmaschinerie besitzen, die an der synaptischen Vesikelexozytose beteiligt ist.
Bei dem Versuch, die mögliche Rolle des SOD1-Exports zu evaluieren, haben wir kürzlich in der Neuroblastom-Zelllinie SK-N-BE gezeigt, dass dieses Enzym durch die Beteiligung des muskarinischen M1-Rezeptors in der Lage ist, ERK1/2 und AKT in einer dosis- und zeitabhängigen Weise zu aktivieren. Dieser Effekt wurde sowohl durch M1-Rezeptor-Silencing als auch durch den M1-Antagonisten Pirenzepin deutlich reduziert (Damiano et al., 2013).
Es wurde jedoch gezeigt, dass FGF-1 und die 18 kDa-Isoform von FGF-2 über einen alternativen Weg sezerniert werden, indem sie direkt aus dem Zytoplasma in den extrazellulären Raum transloziert werden. Analog dazu wurde auch für Interleukin 1β (IL-1β) berichtet, dass es über einen vesikulären, nicht-klassischen Exportweg sezerniert wird. Lösliche Proteine enthalten klassischerweise N-terminale Signalpeptide, die sie zum Translokationsapparat des Endoplasmatischen Retikulums (ER) leiten (Walter et al., 1984).
Darüber hinaus ist die nichtklassische Proteinsekretion sowohl energie- als auch temperaturabhängig und kann durch verschiedene Behandlungen stimuliert oder gehemmt werden (Cleves, 1997; Hughes, 1999). Die Liste der Proteine, die in Abwesenheit eines funktionsfähigen ERG-Systems (unkonventioneller sekretorischer Weg) aus Zellen exportiert werden könnten, wie IL-1β und Galectin-1 (auch als L-14 bezeichnet), wird ständig erweitert; für weitere Daten siehe den Review von Nickel (2003).

Aus diesem Grund sollte die SOD1-Sekretion den kanonischen ERG-Sekretionsweg umgehen. Wir haben zuvor gezeigt, dass BFA, ebenso wie 2-Desoxyglucose und Natriumazid (NaN3), den SOD1-Export beeinträchtigt (Mondola et al., 2003). Unserer Meinung nach dysreguliert die Behandlung mit BFA wahrscheinlich nicht nur den klassischen sekretorischen ERG-Weg, sondern auch den mikrovesikulären Membranverkehr der unkonventionellen Proteinsekretion oder des alternativen Proteinexports.
Der 20-Sekunden-Trick für Superoxid-Dismutase: Gesundheitliche Vorteile, Verwendungen, Nebenwirkungen ...
Außerdem zeigten diese Autoren, dass SOD1A4V den sekretorischen Proteintransport vom ER zum Golgi-Apparat hemmt. Amyotrophe Lateralsklerose (ALS) ist eine im Erwachsenenalter auftretende, neurodegenerative Erkrankung, die durch ein selektives Absterben der oberen und unteren motorischen Neuronen des Gehirns und des Rückenmarks gekennzeichnet ist. Zu den Symptomen gehören Muskelschwund, Spastizität, Lähmung und schließlich Tod durch Atemstillstand innerhalb von 3-5 Jahren nach der Diagnose.
Klinische und pathologische Prozesse weisen darauf hin, dass ER-Stress einen Schlüsselweg darstellt, der am Zelltod beteiligt ist. Im transgenen SOD1G93A-ALS-Rattenmodell wurde eine Unfolded-Protein-Reaktion und ER-Stress-induzierte Apoptose beobachtet (Atkin et al., 2006); eine Unfolded-Protein-Reaktion, einschließlich der Induktion von Stress-Sensor-Kinasen, Chaperonen und apoptotischen Mediatoren, wurde auch in Motorneuronen des Rückenmarks von menschlichen Patienten mit der sporadischen Form der ALS (sALS) gezeigt, die nicht auf SOD1-Mutationen beschränkt ist (Atkin et al., 2008).
SOD1 und andere Proteine sind bei fALS und bei sALS fehlgefaltet, aber es ist nicht klar, wie dies ER-Stress, Fragmentierung des Golgi-Apparats, Störung des axonalen Transports und Apoptose auslöst. Nahezu 20% der fALS wird durch SOD1-Genmutationen verursacht (Neumann et al., 2006). In der Tat behalten die meisten SOD1-Mutanten ihre enzymatische Aktivität bei, was eher auf einen Funktionsgewinn der toxischen Aktivität als auf einen einfachen Funktionsverlust hindeutet (Strong et al., 2005; Dion et al., 2009).
(2005) zeigten eine gestörte konstitutive extrazelluläre Sekretion von mutiertem SOD1 in NSC-34-Zellen, die häufige zytoplasmatische Einschlüsse und Proteinunlöslichkeit induziert. Diese Daten bringen die mangelhafte Sekretion von mutiertem SOD1 mit intrazellulären Proteinaggregaten und Toxizität in NSC-34-Zellen in Verbindung. Darüber hinaus zeigten diese Autoren, dass in einem transgenen Rattenmodell der ALS die chronische intraspinale Infusion von exogenem humanem wt-SOD1 das Fortschreiten der Krankheit signifikant verzögerte, was auf eine neuartige extrazelluläre Rolle von SOD1 bei der ALS hindeutet; daher konnte die extrazelluläre Verabreichung von humanem wt-SOD1 die klinische Erkrankung in transgenen ALS-Ratten verbessern, was eine neuartige extrazelluläre Rolle von mutiertem und wt-SOD1 in der ALS-Pathogenese bzw. Therapie unterstützt.
Darüber hinaus wurden in transgenen Mäusen, die SOD1-Mutationen tragen, toxische Effekte auf motorische Neuronen durch Mikroglia-Aktivierung beobachtet (Urushitani et al., 2006; Zhao et al., 2010). Die Mikrogliazellen können eine wichtige Rolle in der ALS-Progression übernehmen (Pramatarova et al., 2001), da eine Mikrogliaaktivierung vor dem Neuronenverlust in transgenen Mäusen, die humane SOD1-Mutanten exprimieren, beobachtet werden kann (Alexianu et al., 2001).
Vielen Dank für das Teilen Ihrer Informationen. Ich schätze wirklich Ihre Bemühungen und ich warte auf Ihre weitere write ups danke noch einmal. Tandie Lazarus Hughes